- South America
- Central & North America
- Europe & North/Central Asia
- Sub-Saharan Africa
- North Africa & the Middle East
- South Asia
![]()
![]()
![]()
Sub-Saharan AfricaEthiopian wolf Canis simensis

Ethiopian wolf - © Claudio Sillero
ProjectsPublicationsRelevant LinksReports / PapersOther NamesEnglish: Ethiopian Wolf, Simien Fox, Simien Jackal
French: Loup D'Abyssinie
Spanish; Castilian: Lobo Etiope
German: Abessinischer Fuchs, Aethiopienfuchs, Äthiopischer Wolf
Italian: Volpe Rossa
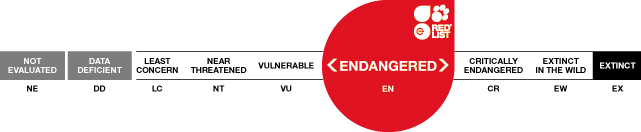
Justification
Notes on population estimates:
Bale minimum estimate of N=210 adults was derived from intensive observations of wolf packs across the Bale Mountains. In April 2009 the population was estimated at 291 adult (>1 year old) wolves (EWCP annual report 2008–2009) and in April 2010 at 290–320 (EWCP annual report 2009-2010). Since then 23 carcasses have been retrieved, and 56 individuals have gone missing, probably as a result of a canine distemper epizootic that affected several subpopulations in Bale.
Ethiopian Wolves are restricted to seven isolated mountain enclaves in the highlands of Ethiopia. The Afroalpine range available to the species is shrinking and increasingly fragmented. The species’ extent of occurrence is estimated at 4,200 km² with an area of occupancy of 2,779 km². Population estimates across the species’ range suggest that between 360 and 440 adult (>1 year old) wolves remain, of which <250 are mature individuals. One relict population in northern Ethiopia (Mt Guna) is now functionally extinct.
The largest population occurs in the Bale Mountains in southern Ethiopia, where intensive monitoring of wolf packs indicate that population size varies widely due to the recurrence of infectious disease epizootics affecting local subpopulations. Since 2008 to date this population has declined by ~30%, from approximately 300 to an estimated 210 adults, due to consecutive epizootics of rabies and canine distemper. This equates to a 20% global decline in the population in the last three years; although narrowly below the threshold for uplisting to Critically Endangered under C1, this continued decline merits careful ongoing monitoring of population numbers (and a reassessment if necessary) and sustained conservation interventions.
Number of "Mature Individuals"
The number of mature individuals in the population (population size) was determined using the IUCN's new definition (in: Guidelines for Using the IUCN Red List Categories and Criteria) and the following formula:
Mature individuals = 0.31N1+ 0.4N1+0.57N2 +0.16 N2 then
Mature individuals = 0.71N1+ 0.73 N2 where:
N1= adult male population
N2= adult female population
Calculations of mature individuals capable of reproduction were derived from empirical data on wolf populations in the Bale Mountains, between 1988–1992 and 1997–2000. Of all adult males in a population, 31% are alpha males and thus reproductive individuals (= 0.31N1). Of all subordinate adult males (= 0.69N1), some may access reproduction either by replacing the alpha male or through extra-pack copulations. This proportion was subjectively estimated at 60% and thus the contribution of subordinate adult males potentially capable of reproduction is 0.6 x 0.69N1 = 0.41N1.
Of all adult females in a population, 57% are in alpha position and thus reproduce (= 0.57N2). Of the suppressed adult females (= 0.33N2) we estimated a 50% likelihood to access reproduction in their lifetime (0.5 x 0.33N2=0.16N2). This was derived from the rate of breeding opportunities, calculated as 0.12 for female per year from empirical data, over four years (0.12 x 4 years= 0.5).
Population estimates were derived from current availability of habitat types and extrapolations of the density of adult and subadult wolves to the area of each habitat type. Population estimates (N) are numbers of adult and subadult wolves. Given a population sex ratio of 2:1 males to females then the male population is 2/3N = 0.67N and the female population is 1/3N= 0.34N. Eighty percent of males and 70% of females are adults, therefore:
N1= adult male population= 0.8 x 0.67N = 0.53N
N2= adult female population= 0.7 x 0.33N = 0.23N
and, Mature individuals = 0.71N1 + 0.73N2, so:
Mature individuals = 0.37N + 0.17N= 0.54N
Mature individual estimates for 2010 based on IUCN/SSC Canid Specialist Group (2011):
Bale Mountains: N = 210; mature individuals = 113
Arsi Mountains: N = 54; mature individuals = 29
Simien Mountains: N = 52; mature individuals = 28
North Wollo: N = 19; mature individuals = 10
South Wollo: N = 16; mature individuals = 9
Menz : N = 15; mature individuals = 8
Mt Guna: N = 0; mature individuals = 0
Total: N = 366; mature individuals = 197
Guassa-Menz estimate of 15 adult wolves was derived from intensive observation of wolf packs in recent years which yielded an estimate of 25 adult wolves, and was corroborated by DNA fingerprinting in 2008 (which identified 23 individuals). During 2010 40% of the known wolves have died or disappeared due to canine distemper, resulting in the revised estimate.
Arsi and Simien estimates (EWCP unpublished) were derived from intensive observations of wolf packs across both ranges. Recent improvements in monitoring resulted in a larger estimate for Simien, and a smaller one for Arsi. These revised figures reflect improved field techniques and effort rather than actual population change.
For North and South Wollo, population estimates are conservative and were based on habitat availability. Using field maps, the extent of suitable habitat were estimated and populations extrapolated from known densities in Bale for habitats with similar characteristics (Marino 2003a). Here the lower value of densities observed were used (i.e., good habitat: 0.2 wolves/km²; marginal habitat 0.1 wolf/km²).
Other parameters used:
Generation time: mean 4.5 yr, range 3–8; extent of occurrence: 4,200 km² [defined as area above 3,000 m asl]; area of occupancy: 2,800 km²; number of populations: 7; number of sub-populations: 10.
Given the information above, Canis simensis meets the criteria for Endangered under criterion B based on <5,000 km² extent of occurrence (B1), as the population is severely fragmented (a), and there is a continuing decline (b) in area, extent and or quality of habitat (iii) and number of mature individuals (v). It also meets the criteria under C based on <2,500 mature individuals in the population (EN C and D), exceeding a 20% decline in the population in the past eight years (two generations), and a continuing decline in population size (EN C2) and <250 mature individuals in each subpopulation (EN C2a(i)). Finally, it also meets the threshold of <250 mature individuals for listing under D.
Geographic Range Information
Endemic to the Ethiopian highlands, above the tree line at about 3,200 m. There are no recent records of the species at altitudes below 3,000 m, although specimens were collected at 2,500 m from Gojjam and north-western Shoa at the beginning of the century (references in Yalden et al. 1980). Reported in the Simien Mountains since the species was first described in 1835, but scattered and irregular sightings suggest numbers have been declining. Reported on the Gojjam plateau until the early 1900s (Powell-Cotton 1902, Maydon 1932). South of the Rift Valley, wolves have been reported in the Arsi Mountains since the 1920s, and, more recently (1959), in the Bale Mountains. Reports of small populations in North Sidamo (Haltenorth and Diller 1980) may be in error. There is no evidence that the Ethiopian Wolf ever occurred in Eritrea (Coetzee 1977).
The species currently is confined to seven isolated mountain ranges of the Ethiopian highlands, at altitudes of 3,000–4,500 m (Gottelli and Sillero-Zubiri 1992, Marino 2003a). In the northern highlands wolves are restricted to land above 3,500–3,800 m by increasing agricultural pressure (Yalden et al. 1980, Marino 2003a). Wolf populations occur north of the Rift Valley in the Simien Mountains, Mount Guna, North Wollo and South Wollo highlands, and Menz. Recently extinct in Gosh Meda (North Shoa), and absent from Mt Choke, Gojjam, for a few decades. South-east of the Rift Valley there are populations in the Arsi Mountains (Mt Kaka, Mt Chilalo and Galama range) and in the Bale Mountains, including the Somkaru-Korduro range (Marino 2003a).
Population trend:Decreasing

Population Information
More than half of the species' population lives in the Bale Mountains, where wolf density is high for a social carnivore of its size, and positively correlated with density of rodent prey and negatively with vegetation height (Sillero-Zubiri and Gottelli 1995). Highest wolf densities are found in short Afroalpine herbaceous communities (1.0–1.2 adults/km²); lower densities are found in Helichrysum dwarf-scrub (0.2/km²), and in ericaceous heathlands and barren peaks (0.1/km²). Wolves are also present at low density (0.1–0.2/km²) in montane grasslands at lower altitudes.
Elsewhere, overall wolf density is relatively lower. In Menz, wolf density was estimated at 0.2 animals/km² using transect data (Ashenafi 2001). Comparison of census transect data from recent comprehensive surveys (Marino 2003a) indicates higher abundance in North Wollo (0.20 ± 0.20 sightings per km), intermediate in Arsi and Guna (0.10 ± 0.11 and 0.10 ± 0.14, respectively), and lower in South Wollo and Simien (0.08 ± 0.13 and 0.06 ± 0.11, respectively). These results were supported by counts of wolf signs (i.e., diggings and droppings) and interview results.
A summary of population abundance and status in each region can be found in Sillero-Zubiri et al. (2004), with the most recent information on population status found in IUCN/SSC Canid Specialist Group (2011), and summarized above. Time series from long-term monitoring of wolves in the Bale Mountains of southern Ethiopia, spanning over 20 years, indicated marked variation in wolf abundance in association to rabies epizootics affecting high-density populations in 1991, 2003 and 2008 (Marino 2003b, Marino et al. 2006, IUCN/SSC Canid Specialist Group 2011). Population numbers returned to previous levels after the first two disruptions, evidencing resilience to catastrophes, but at very low density population growth was unexpectedly low, due to delays in the formation of new breeding packs (Marino et al. in review). Canine distemper (CDV) appears to be more important than previously thought, with CDV outbreaks affecting wolves across subpopulations in the Bale Mountains in 2005, 2006 (Malcolm 2006), and 2010 (Gordon et al. 2010).
Habitat and Ecology Information
A very localized endemic species, confined to isolated pockets of Afroalpine grasslands and heathlands where they prey on Afroalpine rodents. Suitable habitats extend from above treeline at about 3,200 m up to 4,500 m, with some wolves present in montane grasslands at 3,000 m. However, subsistence agriculture extends up to 3,500–3,800 m in many areas, restricting wolves to higher ranges (Marino 2003a). Rainfall at high altitude varies between 1,000 and 2,000 mm/year, with one pronounced dry period from December to February/March.
Wolves utilize all Afroalpine habitats, but prefer open areas with short herbaceous and grassland communities where rodents are most abundant, along flat or gently sloping areas with deep soils and poor drainage in parts. Prime habitats in the Bale Mountains are characterized by short herbs (Alchemilla spp.) and grasses and low vegetation cover, a community maintained in continuous succession as a result of molerat (Tachyoryctes macrocephalus) burrowing activity. Other good habitats include tussock grasslands (Festuca spp., Agrostis spp.), high-altitude scrubs dominated by Helichrysum spp. and short grasslands in shallow soils. In northern parts of the range, plant communities characterized by a matrix of 'guassa' tussock grasses (Festuca spp.), 'cherenfi' bushes (Euryops pinifolius) and giant lobelias (Lobelia rhynchopetalum) sustain high rodent abundance and are preferred by wolves. Ericaceous moorlands (Erica and Phillipia spp.) at 3,200–3,600 m are of marginal value, with open moorlands having patches of herbs and grasses which are relatively good habitat.
Threats Information
Continuous loss of habitat due to high-altitude subsistence agriculture represents the major threat. Sixty percent of all land above 3,200 m has been converted into farmland, and all populations below 3,700 m are particularly vulnerable to further habitat loss, especially if the areas are small and of relatively flat relief (Marino 2003a). Habitat loss is exacerbated by overgrazing of highland pastures by domestic livestock, and in some areas habitat is threatened by proposed development of commercial sheep farms and roads. Human persecution triggered by political instability in the past is currently less severe and associated with conflicts over livestock losses (Marino 2003a). Recent population decline in Bale has been mostly due to disease epizootics, with road kills and shooting as secondary threats. Rabies is a potential threat to all populations, with canine distemper also a concern in Bale. Most of these threats are exacerbated by the wolves' specialization to life in the Afroalpine ecosystem.
In Bale, the Ethiopian wolf hybridizes with domestic dogs (Canis familiaris). Gottelli et al. (1994) used mitochondrial DNA restriction fragments and microsatellite alleles to conclude that hybridization was relatively common in western Bale as a result of crosses between female wolves and male domestic dogs. Hybrids have shorter muzzles, heavier-built bodies and different coat patterns. Although hybrids are confined to the Web Valley in western Bale they may threaten the genetic integrity of the wolf population. Following hybridization, a population may be affected by outbreeding depression or reduction in fitness, although to date this does not seem to have taken place in Bale. Currently there is no indication of hybridization taking place outside western Bale.
There is no exploitation of the species for furs, body parts or other purposes.
Use and Trade Information
Conservation Actions Information
The species is not included on the CITES Appendices. It has full official protection under Ethiopia's Wildlife Conservation Regulations of 1974, Schedule VI. Killing a wolf carries a sentence of up to two years.
The Ethiopian Wolf occurs in several protected areas: Bale Mountains National Park; Simien Mountains National Park; Borena Saiynt Regional Park (South Wollo); Guassa Community Conservation Area (North Shoa); Arsi Mountains Regional Park. As a result of boundary extensions (Simien) and new parks created (Arsi) the area of suitable wolf habitat within protected areas increased to 87%.
A number of important steps have been taken in the interests of conserving this endemic species, including: 1) a dog vaccination campaign in Bale, and vaccination campaigns in response to rabies outbreaks in dogs close to wolves in Menz and Simien; 2) sterilization programme for hybrids in Bale; 3) vaccination of wolves in response to rabies epizootics in parts of Bale; 4) community and school education programme in Bale and Wollo; 5) strengthening the capacity of the Bale Mountains National Park – funding patrolling, maintenance of infrastructure, etc.; 6) surveys to determine the persistence and status of all populations of wolves; 7) monitoring of all wolf populations; 8) A Strategic Planning workshop for Ethiopian wolf conservation held in Lalibela, February 2011, resulting in a National Action Plan for the next 10 years (IUCN/SSC Canid specialist Group 2011).
In 1983, the Wildlife Conservation Society established the Bale Mountains Research Project, which publicized the wolf's plight and started a regular monitoring programme for the species. A detailed four-year field study followed (Sillero-Zubiri 1994). Based on its findings, the IUCN/SSC Canid Specialist Group produced an action plan for the Ethiopian Wolf (Sillero-Zubiri and Macdonald 1997), which provided a detailed strategy for the conservation and management of remaining wolf populations. This plan advocated immediate action on three fronts – education, wolf population monitoring, and rabies control in domestic dogs – to conserve the Afroalpine ecosystem and its top predator. As a result, the Ethiopian Wolf Conservation Programme (EWCP) was established in 1995 by Oxford University with financial support from the Born Free Foundation, Frankfurt Zoological Society (FZS), the Wildlife Conservation Network and other donors. The Programme’s overall aim is to protect the Afroalpine ecosystem and many of its rare highland endemic plants and animals through better management in Bale and the establishment of other conservation areas in Menz and Wollo. The EWCP currently monitors the demography of the Bale and selected populations in South and North Wollo, supports park patrols within the wolf range, undertakes domestic dog control and the removal of dog-wolf hybrids. Additionally, the EWCP carries out a community conservation education campaign that targets people living inside the wolf's range and is aimed at improving dog husbandry and combating disease in the park and surroundings. A large-scale dog vaccination programme (targeting up to 5,000 dogs a year) seeks to reduce the occurrence of rabies and distemper within the Ethiopian wolf range and is backed up by further epidemiological and demographic studies. The EWCP is also active elsewhere in Ethiopia, with representatives surveying and monitoring all wolf ranges and implementing education campaigns about the plight of the species. Zelealem Tefera Ashenafi set up the Guassa Biodiversity Project in 1996, looking at the relationships between pastoralists and wildlife in the highlands of Menz. FZS and partners are working in North Ethiopia to protect the Afrolapine ecosystem, supporting the Simien Mountains National Park and creation of a new community conservation area in North Wollo.
There are no animals maintained in captivity.
Although the behavioural ecology of the species is well known, research has focused largely in the optimal habitats in the Bale Mountains. Additional information on dispersal distance and survival would be useful. Investigation into the role of the species in the epidemiology of canid-related diseases is necessary. Studies on wolf-prey relationships and prey availability in the high risk populations of northern Ethiopia are also needed.

