- South America
- Central & North America
- Europe & North/Central Asia
- Sub-Saharan Africa
- North Africa & the Middle East
- South Asia
![]()
![]()
![]()
Central & North AmericaIsland fox Urocyon littoralis

island fox © Coonan
Island, Kit & Swift Fox Working Group - Brian Cypher and Donelle Schwalm are the coordinators of the Island, Kit & Swift Fox Working Group. The working group report that for both kit foxes and swift foxes, habitat continues to be lost and remaining habitat is increasing subject to degradation, fragmentation, and incompatible uses. The future prognosis for island foxes is however significantly improved. After 4 of 6 subspecies suffered catastrophic declines due to novel predators and disease, mitigation measures have been immensely successful and these 4 populations have returned to or are approaching pre-decline levels. This page highlights some of the recent issues and current conservation needs for these species.
Relevant Links- Flickr: Wild Canids of the World
- 2013 IUCN Red List Assessment - Island Fox
- National Park Service - Island Fox
- Institute for Wildlife Studies - Island Fox
- Friends of the Island Fox
- Catalina Island Conservancy
- The Nature Conservancy - Santa Cruz Island Fox
- Defenders of Wildlife - Channel Island Fox
Generic synonym: Vulpes
English Island Fox, California Channel Island Fox, Channel Islands Fox, Island Gray Fox, Island Grey Fox
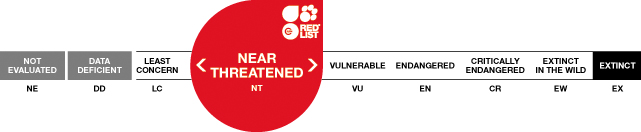
Justification
The Island Fox is restricted to six of the California Channel Islands off the coast of southern California, USA. Four Island Fox subspecies suffered catastrophic population declines in the mid-1990s, after which they were listed as Endangered under the U.S Endangered Species Act. However, all four subspecies have since recovered to or are approaching recovery to pre-decline population levels, due to the success of aggressive recovery actions such as captive breeding and reintroduction, relocation of Golden Eagles, and vaccination against canine diseases. The total population of mature individuals increased from <1,500 in 2002 to >4,000 in 2011 and annual survival rates are above 85%.
The Red List Status for the Island Fox is currently assessed as Near Threatened (it almost qualifies as threatened under criteria B1a and C2a). The species meets none of the criteria for the threatened categories (Critically Endangered, Endangered or Vulnerable). In regard to criterion A, population reduction, the species has not experienced a population reduction in the past 10 years or three generations. Four subspecies experienced declines in the 1990s, but have since increased significantly, to near pre-decline numbers. Criterion B concerns geographic range. The total range for the species is about 9,000 km², and the number of locations (considering past and future potential threats) is <10 (the species occurs on six islands) which would place the species in the Vulnerable category, but only if there was a continuing decline or extreme fluctuation in occurrence, occupancy, locations, or population size. Criterion C considers small population size. The number of mature individuals is approximately 4,000, whih would place the species in the Vulnerable category, but only if there was also a continuing decline and the number of mature individuals was <1,000 in each subpopulation, or if the population fluctuated extremely . Island Foxes do not meet criterion D, very small or restricted population size, because the number of mature individuals is >1,000 and number of locations is >5. Finally, Island Foxes do not meet the criteria for E, quantitative analysis. Recent estimates indicate that the probability of extinction in the next 50 years is <5% for each of the four Island Fox subspecies listed as endangered under the US Endangered Species Act (T. Coonan, National Park Service, C. Boser, The Nature Conservancy and J. King, Santa Catalina Island Conservancy, unpubl. data).
Although the species meets none of the criteria for the threatened categories, it is assessed as Near Threatened because of inherently small population sizes (none of the six subspecies exists at adult population levels considerably higher than 1,000 individuals), potential threats from disease and/or predation, and the continuing need for intensive management actions such as population size and survival rate monitoring, disease and Golden Eagle surveillance, and annual vaccination against rabies and canine distemper virus. If considered alone, some of the subspecies would likely meet criteria for a threatened category, due primarily to inherently small population sizes. However, previous Red List assessments have been conducted at the species level for Island Foxes. Because recent threats to subspecies have been mitigated, it is not necessary to consider Red List status for individual subspecies at this time.
According to IUCN Red List Guidelines (IUCN Standards and Petitions Subcommitee 2013), a species may be moved from a category of higher threat to one of lower threat if none of the criteria of the higher threat category has been met for five years or more. Therefore, to be moved to the Near Threatened category in 2013, Island Foxes must not have met criteria from any higher threat category (Critically Endangered, Endangered or Vulnerable) criteria in 2008 or later. The following demonstrates, category by category, that Island Foxes did not meet any of the threatened category criteria in 2008.
A. Population reduction:
As of 2008, had Island Foxes undergone a population decline over either 10 years or three generations (whichever is longer)? A recent life table matrix model (B. Hudgens, Institute for Wildlife Studies, unpubl. data) indicates Island Fox generation time is about four years. Therefore, from 1996-2008, was there a 90%, 70% or 50% decline in the overall number of island foxes? By 2008 the number of island foxes was approximately 2,500 adults, but the population in 1996 is unknown. In that time period, 1996-2008, four Island Fox subspecies had reached the nadir of their declines (in 1999-2000) and recovery was well under way on all islands, so much so that captive breeding had ceased on all islands by 2008. Thus, it is unlikely that there was a 50% or greater decline between 1996 and 2008, a period spanning three generations of Island Foxes.
B. Geographic range:
The range of Island Foxes comprises about 9,000 km², which meets the Vulnerable category if two of the following are met: the number of locations is less than 10, a continuing population decline, or extreme fluctuations. Although Island Foxes occur in <10 locations (six islands), there was no continuing decline in 2008, nor were there extreme fluctuations. Therefore, Island Foxes do not meet the geographic range criteria for any of the threatened categories.
C. Small population size and decline:
The number of mature Island Foxes was about 2,500 in 2008. That would place the species in the Endangered category, but only if there was a continuing decline, or a decline combined with either a) a low mature population size in each subpopulation, b) a high proportion of individuals occurring in just one subpopulation or c) or extreme fluctuations. Because there was no continuing decline in 2008, Island Foxes do not meet the small population size and decline criteria for any of the threatened categories.
D. Very small or restricted population:
The number of mature individuals in 2008 was 2,500, which is above the 1,000 required for a Vulnerable rating. Additionally, they did NOT have an area of occupancy of <20 km² or number of locations <6. Therefore, Island Foxes do not meet the very small or restricted population criteria for any of the threatened categories.
E. Quantitative Analysis:
There is no analysis that concludes that in 2008 the probability of extinction for Island Foxes was 50% in 10 years (Critically Endangered), 20% in 20 years (Endangered) of 10% in 100 years (Vulnerable). Thus, Island Foxes do not meet the quantitative analysis criteria for any of the threatened categories
Geographic Range Information
Island Foxes are restricted to the six largest of the eight California Channel Islands located off the coast of southern California, USA. The view that foxes reached the northern Channel Islands by waif dispersal, followed by Native American assisted dispersal to the southern Channel Islands (Collins 1982, 1991a,b, 1993; Wayne et al. 1991; Goldstein et al. 1999), was challenged by Rick et al. (2009), who dated fossil fox bones. The oldest bone was dated at approximately 6,400 cal yr BP, several thousand years after humans colonized the islands. Thus, it is now unclear whether foxes reached the northern islands naturally or were introduced by humans.
Six distinct subspecies are recognized, one on each of the islands where they occur:
San Miguel Island Fox (Urocyon littoralis littoralis (Baird, 1858)), San Miguel Island,
Santa Rosa Island Fox (U. l. santarosae Grinnell & Linsdale, 1930), Santa Rosa Island,
Santa Cruz Island Fox (U. l. santacruzae Merriam, 1903), Santa Cruz Island,
Santa Catalina Island Fox (U. l. catalinae Merriam, 1903), Santa Caralina Island,
San Nicolas Island Fox (U. l. dickeyi Grinnell & Linsdale, 1930), San Nicolas Island, and
San Clemente Island Fox (U. l. clementae Merriam, 1903), San Clemente Island.
Population trend:Increasing

Population Information
Island Foxes exhibit substantial variability in abundance, both spatially and temporally. In the 1990s, Island Fox numbers declined on five of the six islands where they occur, reducing the total population from approximately 6,000 individuals (Roemer et al. 1994) to less than 1,500 in 2002. Island Fox numbers have been steadily increasing in the past decade, reaching a total population of approximately 5,500 and an adult population of >4,000 in 2011.
Fox populations on both San Miguel and Santa Cruz Islands declined by > 90% between 1995 and 2000. Similar declines also occurred on Santa Rosa and Santa Catalina Islands (Roemer 1999; Timm et al. 2000; Roemer et al. 2001a, 2002; Coonan 2003). By 2000, Island Fox populations on San Miguel and Santa Rosa islands had declined to only 15 individuals, all of which were brought into captivity (Coonan 2002, 2003; Coonan et al. 2010a). The Santa Cruz population dropped from an estimated 1,300 foxes in 1993 to as low as 50 foxes in 2001, including 10 pairs brought into captivity in 2002 (Roemer 1999, Roemer et al. 2001, Coonan et al. 2010a). Captive populations were maintained on the islands from which they were originally captured. Beginning in 2003, captive-bred foxes from each of the three northern islands (San Miguel, Santa Cruz and Santa Rosa) were released back into the wild. By 2008, all captive animals were released and populations were growing exponentially on all three islands. By 2011, wild fox populations had increased to an estimated 580 foxes with 400 adults on San Miguel Island, 450 foxes with 280 adults on Santa Rosa Island, and 1,200 foxes with 880 adults on Santa Cruz Island (Coonan 2012).
On Santa Catalina, Island Foxes were nearly completely eliminated on the eastern portion of the island by a canine distemper outbreak that swept through the population in 1999 (Timm et al. 2000), with only about 110 adult foxes remaining on the island in 2000. Translocations from the western portion of the island, beginning in 2003, have resulted in a steadily growing population, estimated at approximately 1,500 foxes in 2011. About 80% of the fox population now lives on the island’s east end.
The San Clemente population has steadily increased from approximately 250 adults in 2001-2002 (Wolstenholme et al. 2003 ) to approximately 700 adults in 2011 (Gregory et al. 2012), with a total population (including young of the year) estimated around 1,100 (Garcia and Associates 2011). A small area of stabilized sand dunes on the north end of the island support the highest recorded density of foxes with density estimates ranging from 14--40 foxes/km² (Gregory et al. 2012, Sanchez and Hudgens 2011). Outside of the dunes, fox densities ranged from 2.5-7.8 adults/km² with densities generally highest in the north and lowest in the south (Gregory et al. 2012).
In contrast to the other populations, the San Nicolas Island Fox population has remained relatively stable over the past 10 years, though there is anecdotal and genetic evidence of a severe population decline to an effective population size of less than 10 breeding individuals in the 1970s (Laughrin 1980, Aguilar et al. 2004). Since 2000, estimated population size has fluctuated from 500-700 adults and 600-800 total, including young of the year (Garcelon and Hudgens 2012). The highest densities of animals occur in the stabilized dunes and coastal sage scrub on the western end of the island, where adult fox densities of over 25 foxes/km² have been recorded. Fox densities are much lower in the grasslands and barren cliffs in the south and east of the island, where recorded densities are only 1.9 foxes/km² (Garcelon 2010 ).
All current estimates of density and population size in island foxes have been derived from trapping data using modifications of a capture-recapture approach (Roemer et al. 1994). Density estimates prior to 2005 were generally calculated by dividing estimated population size within a trapping grid by the effective trapping area including a buffer determined by the mean-maximum distance moved (MMDM) by captured individuals. More recent density estimates were generally estimated from spatially-explicit mark-recapture analyses (Efford et al. 2004). Island-wide estimates were extrapolated either by simply multiplying the average densities of trapped areas by the total island area, or by multiplying densities of trapped areas within different habitat types by the area of each habitat on the island.
Habitat and Ecology Information
The Island Fox is considered to be a habitat generalist because it has been found using virtually all habitat types occurring on the Channel Islands (Coonan et al. 2010a, USFWS 2012). Use of diverse habitats likely is a function of ecological plasticity, limited predation and competition pressures, and characteristic high fox densities. Habitats used include beaches, sand dunes, bluffs, grasslands, sage scrub, cactus scrub, chaparral, oak woodlands, riparian woodlands, pine forests, marshes, and developed areas (e.g., towns, campgrounds). However, preferential use of some habitats has been detected. Island Foxes may prefer stabilized sand dunes (Sanchez and Hudgens 2012, Gregory et al. 2012, Garcelon and Hudgens 2012) and areas with higher topographic and vegetation structure diversity (USFWS 2012) such as shrublands and woodlands (Laughrin 1977, Crooks and Van Vuren 1996, Roemer 1999, Drake 2013). Conversely, some habitats appear to be used less, particularly types with low vegetation structure such as grasslands (Roemer 1999, Roemer and Wayne 2003).
Home range use by Island Foxes is influenced by various factors including topographic features, available habitat types, habitat-specific resource abundance, fox density, sex, and age (USFWS 2012). Mean home range sizes of 0.16-3.39 km² have been reported (Coonan et al. 2010a, Sanchez 2012, Drake 2013) Size generally is inversely related to fox density (Sanchez 2012) and is slightly larger for males. Also, foxes using grasslands tend to have larger home ranges than foxes using denser, more structurally diverse habitats. Den sites are variable and can occur in rocks, brush, log piles, earthen burrows, and man-made structures (Laughrin 1977, Moore and Collins 1995). Island Foxes exhibit both nocturnal and diurnal activity, and also are skilled climbers thereby allowing them to access resources in trees and large shrubs (Coonan et al. 2010a).
Island Foxes are foraging generalists and consume a wide variety of items including rodents, birds, lizards, insects, snails, carrion, and fruits (Moore and Collins 1995, Cypher et al. 2011). Use of items varies by habitat-specific and island-specific availability, and also varies by seasonal availability (e.g., fruits, some insects, marine mammal carrion). Preferred items appear to include deer mice (Peromyscus maniculatus), lizards, beetles, Jerusalem crickets (Stenopalmatus spp.), earwigs (Forficula auricularia), and fruits of toyon (Heteromeles arbutifolia), manzanita (Arctostaphylos spp.), and prickly pear cactus (Opuntia spp.) (Cypher et al. 2011). Deer mice may be a particularly important food for parents to feed to growing pups due to ease of transport and high protein content (Laughrin 1977, Garcelon et al. 1999).
Threats Information
Predation by non-native Golden Eagles (Aquila chrysaetos) led to a catastrophic decline on the three northern islands (Santa Cruz, Santa Rosa, and San Miguel) during the 1990s but aggressive management actions have greatly reduced this threat (see Conservation section) (Coonan et al. 2010a). A catastrophic decline on Santa Catalina Island was due to canine distemper virus, probably introduced by a raccoon (Procyon lotor) that “stowed-away” on a boat from the mainland (Garcelon et al. 1992, Roemer 1999, Timm et al. 2000, Coonan et al. 2010a). Efforts to control fox predation on San Clemente Island Shrikes, Lanius ludovicianus anthonyi), were previously considered a possible threat to the foxes (Roemer and Wayne 2003) but have been discontinued.
Island Foxes have naturally low population levels (less than 1,500 on the larger islands and less than 500 on San Miguel) (Coonan et al. 2010a). Furthermore, they have extremely low levels of genetic variability (Gilbert et al. 1990, Wayne et al. 1991, Goldstein et al. 1999, Aguilar et al. 2004), and thus reduced ability to adapt to future environmental change. The possible introduction of new diseases from the mainland remains a threat to populations on all islands. Thus, these foxes will always be a “conservation reliant” species, requiring careful monitoring and rapid management interventions to survive (Scott et al. 2010).
Use and Trade Information
Conservation Actions Information
The species is not included in the CITES Appendices.
Legal status
In 2004, four of the six Island Fox subspecies were federally listed as endangered by the U.S. Fish and Wildlife Service (USFWS) under the federal Endangered Species Act (USFWS 2004). The northern Island Fox subspecies (San Miguel, Santa Rosa and Santa Cruz) were listed due to population declines caused by Golden Eagle predation, and Santa Catalina Island Foxes were listed because of a population decline likely due to canine distemper virus. The San Nicolas and San Clemente subspecies were not listed as endangered by USFWS, because USFWS had been petitioned only to list the three northern subspecies and the one on Santa Catalina Island (USFWS 2004). All six subspecies had been previously listed as threatened by the state of California (California Department of Fish and Game 1987).
Need for recovery actions
Northern Channel Islands
Dramatic fox population declines on San Miguel, Santa Rosa and Santa Cruz Islands were detected during the 1990s. The Island Fox population on San Miguel declined from an estimated 450 adults in 1994 to 15 in 1999 (Coonan et al. 2005). The Santa Cruz population declined from as many as 2,000 adults in 1994 to 50–60 in 2000 (Coonan et al. 2010a). Foxes on Santa Rosa may have numbered more than 1,500 in 1994 (Roemer et al. 1994) but declined to 15 animals by 2000 (Coonan and Rutz 2002). Prior to implementation of Island Fox recovery efforts, Roemer (1999) estimated time to extinction at five years for Island Foxes on San Miguel and 12 years for Island Foxes on Santa Cruz.Evidence from radio-telemetry studies showed that predation by Golden Eagles was the primary mortality factor for Island Foxes on the northern Channel Islands, and caused the massive decline of the three northern subspecies from 1994 to 2000 (Roemer et al. 2001). Golden eagle predation was identified as the cause of death for 19 of 21 radio-collared Island Foxes on Santa Cruz Island from 1993 to 1995 (Roemer et al. 2001). On San Miguel Island in 1998–1999, four of eight radio-collared Island Foxes were killed by Golden Eagles in a 4-month period (Coonan et al. 2005). Until the 1990s, Golden Eagles, which prey mainly on small mammals, had never bred on the Channel Islands, and their recent colonization of the islands was due to a prey base, feral pigs (Sus scrofa) and mule deer (Odocoileus hemionus), that was not present prehistorically (Latta et al. 2005; Collins and Latta 2006). The absence of Bald Eagles (Haliaeetus leucocephalus), which bred historically on the islands and prey mainly on fish and seabirds and whose presence may have kept Golden Eagles away, may also have allowed Golden Eagle colonization of the islands (Roemer et al. 2001). Island Foxes evolved in the absence of significant diurnal aerial predators such as Golden Eagles, and therefore may have been more vulnerable to predation than other small carnivores. Moreover, on much of the northern Channel Islands, historic sheep grazing changed the predominant vegetation from shrub to non-native grasslands, which offer much less cover from aerial predators.
Santa Catalina Island
At the same time that the National Park Service (NPS) was dealing with the issue of eagle predation and very low fox populations, The Santa Catalina Conservancy and its contractor, Institute for Wildlife Studies, were addressing a massive population decline of Island Foxes in 1999-2000, apparently caused by canine distemper virus (CDV). Foxes had declined on the eastern portion of the island, but not on the western portion, which was separated from the western portion by a narrow isthmus. The presence of CDV antibodies in Island Fox blood samples, observation of disoriented foxes, and the presence of CDV in a fox carcass led to the conclusion that the rapid decline (within one year) had been most likely caused by an outbreak of CDV. Later analysis indicated that the CDV was most like the strain present in raccoons, suggesting that the source of the diseases was not the unvaccinated dog and cat population on the island, but perhaps a stowaway raccoon (Coonan et al. 2010a).
Recovery plan
The U.S. Fish and Wildlife Service issued a draft recovery plan for the four listed subspecies in 2012 (USFWS 2012). The plan presented recovery criteria that were based on population risk and on threats. To meet population risk-based criteria, fox populations must demonstrate demographic characteristics consistent with long-term viability, as measured by a population recovery tool that estimates extinction risk using adult mortality and population size estimates (Bakker and Doak 2009). To meet threat-based recovery criteria, land managers must develop monitoring programs and plans for mitigating the two primary threats to Island Foxes, disease and predation. The plan’s recovery actions included removing the threats of predation and disease, managing captive island fox populations for recovery, and establishing island fox monitoring strategies. In reality most of those recovery actions had been completed by the land management agencies prior to release of the recovery plan.
Development of recovery actions
The major recovery actions for Island Foxes were those recommended by an ad hoc group of scientists first convened in 1999 by the NPS to recommend emergency actions that would prevent the extinction of Island Foxes (Coonan et al. 2010a). From 1999–2012, the NPS annually convened a group of experts to help evaluate the status of Island Foxes on Park lands, and to make findings regarding appropriate recovery actions. The Island Fox Conservation Working Group comprised a loose affiliation of public agency representatives, landowners, conservancies, zoological institutions, non-profits, and academics concerned about conservation efforts for the Island Fox. The working group served as a forum for information exchange and evaluation of recovery efforts, dividing into subject matter groups to tackle most issues. In most years the subgroups were concerned with captive breeding, wild fox populations, fox health (veterinary) issues, and education. The group annually reported the status of Island Foxes on all islands, listed findings in regard to threats to the species, and recommended appropriate mitigation actions. From 2004-2006 USFWS used the Island Fox Conservation Working Group to develop its draft Island Fox recovery plan.
In 2003, the Park completed a recovery strategy for Island Foxes on the northern Channel Islands (Coonan 2003). The recovery strategy focused on the major recovery actions of Golden Eagle translocation, Island Fox captive breeding and reintroduction, and disease mitigation via vaccination, as well as the larger, ecosystem-level actions of non-native ungulate removal and reintroduction of Bald Eagles. On Santa Catalina Island, the CDV-caused population decline was addressed by the Santa Catalina Island Conservancy and the Institute for Wildlife Studies. At the Conservancy’s request, IWS developed recovery actions, which included development of a vaccine against CDV, development of a captive breeding program, and translocation of foxes from the still populous western portion to the sparsely populated eastern portion of the island (Coonan et al. 2010a).
Implementation of recovery actions
Since 1999, recovery actions have been implemented, and most of them completed, on the northern Channel Islands and on Santa Catalina Island, and conservation measures have been implemented on the two other islands with foxes (San Nicolas and San Clemente). These actions included captive breeding and reintroduction of Island Foxes; capture and translocation to the mainland of Golden Eagles; development of a vaccination program for wild Island Foxes; development of epidemic response plans; removal of non-native ungulates from the northern Channels Islands; reintroduction of Bald Eagles; and development and implementation of Island Fox monitoring programs.
Captive breeding
Development of captive breeding programs occurred simultaneously on the northern Channel Islands (for San Miguel, Santa Rosa and Santa Cruz Island Foxes) and on Santa Catalina Island (Coonan et al. 2010a). Upon receiving recommendations from the Island Fox Conservation Working Group, the Park began taking emergency recovery actions in 1999, focusing on two measures, the removal of the existing Golden Eagles on the islands, and captive breeding of the critically low Island Fox populations.
Because Island Foxes had not been bred in captivity before, the Park worked with local zoo staff, the Association of Zoos and Aquariums and other national experts to establish animal husbandry methods, including facility design and construction, diet, genetic management, pairing and breeding and release techniques (Coonan et al. 2010a, 2010b). Captive breeding was begun in situ on each island, because each subspecies was managed individually, and mainland facilities would have exposed captive foxes to a suite of pathogens unknown on the islands. Captive breeding began on San Miguel in 1999 and on Santa Rosa in 2000, when the remaining 15 foxes on each island were brought into captivity, respectively. In 2001 the IFCWG recommended beginning captive breeding on Santa Cruz, to avoid starting captive breeding with the kind of perilously low numbers that the programs on San Miguel and Santa Rosa began with. Accordingly, 20 foxes were brought into captivity on Santa Cruz in 2002. On Santa Catalina Island, IWS built pens and brought 12 pairs of foxes from the populated west end into captivity in 2001.
Despite the low number of founders, Island Fox captive populations grew quickly, and reached captive facility capacity, and the level at which releases could begin, in 2001 on Santa Catalina, 2003 on Santa Rosa and Santa Cruz, and 2004 on San Miguel. Island Fox captive breeding encountered challenges such as low reproductive success later in the program, evidence of stress in captive foxes, male aggression, female abandonment of pups, and an outbreak of mastitis in the Santa Rosa and San Miguel facilities. Video monitoring of captive foxes allowed identification of factors that led to successful reproduction and rearing (Calkins et al. 2013). Releases ceased in 2004 on Santa Catalina, and 2007-2008 on the northern Channel Islands. On the northern islands, reproductive success in the wild had begun to outpace that in captivity, and so all foxes were released by 2008. There are now no Island Foxes in captivity. Over the captive breeding period, 254 foxes were released to the wild on the northern islands and 57 on Santa Catalina, where reintroduction was augmented by translocation of juvenile foxes from the populous West end to the East end.
Reintroduction was very successful, due to generally high survival and high reproductive success of released foxes (Coonan and Schwemm 2009). Wild populations grew rapidly on San Miguel, Santa Cruz and Santa Catalina, though on Santa Rosa fox survival, and thus population growth, was retarded by Golden Eagle predation (Coonan et al. 2010a). Annual rate of increase (lambda) for recovering fox populations ranged from 1.3-1.9, and by 2011-2012, the adult populations on San Miguel, Santa Rosa and Santa Cruz were estimated at 400, 500, and >900 individuals, respectively, and there were >1500 total foxes on Santa Catalina (Coonan 2012). Foxes had reached pre-decline levels on San Miguel and Santa Cruz, 12 years after declines brought them perilously close to extinction.
Development of Vaccines
Staff from IWS worked with the Smithsonian National Zoological Park on the use of a vaccine against CDV for Island Foxes. The Canary pox virus recombinant CDV vaccine was tested in a small sample of Island Foxes and proved safe and effective, with vaccinated foxes seroconverting and exhibiting positive antibody titers. The vaccine was thereafter administered to all captive foxes on all islands and later to a portion of the wild populations, upon the advice of the Island Fox health group (Coonan et al. 2010a, Coonan 2012). Although the effectiveness of the vaccine has not been tested in a challenge study, evaluation indicates vaccination does produce antibody response consistent with protection, though annual booster is required to maintain adequate antibodies (Coonan et al. 2012). Vaccination of Island Foxes against CDV will continue indefinitely.
Because there is a naturally-occurring morbillivirus in Island Fox populations that likely confers some immunity against CDV (Clifford et al. 2006), not all wild foxes are vaccinated, to allow circulation of the morbillivirus. On most islands, foxes are vaccinated using a “vaccinated core” strategy, in which 80-100 foxes are vaccinated in one or two geographic core areas. This allows the natural morbillivirus to circulate among foxes on the rest of the island; protects foxes in areas that may be access points for CDV (such as near harbors) and theoretically protects a core group of individuals that could be used to start a captive breeding program, should CDV decimate the remainder of the population. On Santa Catalina, however, foxes are vaccinated in an island-wide strategy, because of the large number of landing sites on that island.
Wild Island Fox populations are also currently vaccinated against rabies, because a rabies epidemic among foxes could not be adequately mitigated, once detected. Unlike canine distemper virus, there is no naturally-occurring rabies among island foxes, so the vaccinated core strategy is not implemented; all trapped foxes are vaccinated against rabies (Coonan 2012).
Golden Eagle Removal
The Park established a cooperative agreement with the Santa Cruz Predatory Bird Research Group (SCPBRG) in 1999 for the purpose of relocating Golden Eagles from the northern Channel Islands. Personnel from the SCPBRG began eagle surveys and removal on Santa Cruz Island, the island with the most recent sightings, in late summer 1999. Golden Eagles were discovered breeding on both Santa Cruz and Santa Rosa Islands. By the end of 2006, 44 Golden Eagles had been removed, mostly from Santa Cruz Island, the majority by bownet trapping. Captured birds were released in north-eastern California, and satellite telemetry on the first released birds indicated that none attempted to return to the islands (Latta et al. 2005). Island Fox survival increased as the number of eagles on the island declined (Coonan et al. 2010a).
Genetic analysis of blood samples from captured eagles suggested that initial colonization of the islands by Golden Eagles had been rapid, perhaps the result of one colonization event by a family group (Sonsthagen et al. 2012). Rapid growth of the island eagle population ensued, to the point where carrying capacity was reached and young eagles began dispersing from the island population to the mainland. This implies that preventing future recolonization of the islands by dispersing mainland eagles will be crucial to Island Fox persistence. Actions that will prevent colonization include rigorous monitoring for evidence of predation and eagle presence, removal of non-native eagle prey, and re-establishement of a robust breeding population of territorial Bald Eagles.
Larger ecosystem recovery actions
The team of experts first convened by NPS in 1999 concluded that mitigating Golden Eagle predation on Island Foxes required not only the immediate, emergency recovery actions of eagle relocation and Island Fox captive breeding, but also larger, ecosystem-level actions that would tip the balance away from continued Golden Eagle use of the islands and toward conditions that would allow foxes to persist. Accordingly, the team called for both removal of feral pigs from Santa Cruz Island, since the non-native pigs facilitated the “hyperpredation” visited upon foxes (Roemer et al. 2001), and the reintroduction of Bald Eagles to the northern Channel Islands, positing that the presence of breeding Bald Eagles would discourage nesting by Golden Eagles (Coonan and Rutz 2001). With the later discovery of deer bones in Golden Eagle nests on Santa Rosa Island (Latta et al. 2005; Collins and Latta 2006), the need to remove that prey source became apparent (Coonan and Dennis 2007), and the USFWS Island Fox recovery plan called for the removal of non-native ungulates from that island (USFWS 2012).
Reintroduction of Bald Eagles
The restoration of Bald Eagles to the northern Channel Islands, funded by the Montrose Settlements Restoration Program, comprised annual releases of young eagles from 2002-2006. Breeding by released Bald Eagles began in 2006, and as of 2012 there were >40 bald eagles on the northern Channel Islands, with breeding occurring on Anacapa, Santa Cruz and Santa Rosa islands (Sharpe 2012).
Removal of non-native ungulates
Prior to the discovery that feral pigs supported Golden Eagles on Santa Cruz Island and so facilitated predation on Island Foxes, the NPS had sought to remove feral pigs from that island because of the impacts to natural and cultural resources from pig rooting and foraging (NPS 1985). Accordingly, in 2005-2006 the NPS and The Nature Conservancy, co-owners of Santa Cruz Island, cooperated to remove feral pigs (>4,000 total) from Santa Cruz Island, thereby eliminating that non-native prey source for Golden Eagles (Morrison et al. 2007, Coonan et al. 2010a).
Non-native ungulates had existed on Santa Rosa Island since the early 20th century, when they were bought to the island for hunting purposes. Their phased removal was scheduled as part of a court settlement among the NPS, USFWS, the former island owners, and environmental groups to address ungulate impacts to water quality and endangered plant species (Coonan et al. 2010a). By the end of 2012, all elk had been removed from the island, and only a handful of deer remained (T. Coonan, NPS, unpubl. data).
Monitoring
Thorough Island Fox monitoring programs have been implemented on all six islands. For the four islands with listed subspecies, monitoring programs were established to track effectiveness of recovery actions and to chart the recovery of populations. These initial monitoring efforts have evolved into longer-term programs designed to track Island Fox population size, survival, mortality sources, and exposure to disease.
Because reintroduction and/or translocation occurred on all four of islands with listed subspecies, released foxes were radio-collared to monitor their survival and identify mortality sources. As of 2013, a sample of radio-collared foxes was maintained on all six islands to detect mortality causes such as disease. Annual survival of Island Foxes has been tracked for listed subspecies, and shows increased survival over time, with recovered populations exhibiting annual survival of 80-90% (Coonan et al. 2010a).
On the four islands with listed subspecies, a portion of the 50 or so radio-collared foxes are disease sentinels. These are young, unvaccinated foxes whose mortality may detect emerging diseases as well as the presence of diseases against which a portion of each Island Fox population is vaccinated (rabies and CDV).
For naturally small populations such as those of the Island Fox, population size is important to track, both to chart recovery of populations and to assess demographic viability. Prior to the declines of the 1990s, the NPS tracked population trends of Island Foxes on San Miguel, as part of its longterm ecological monitoring program. Population size of the four endangered subspecies has been monitored since reintroductions or translocations began, and population trends on San Clemente are also monitored. Guidance for population monitoring of all fox populations was developed through the efforts of the Island Fox working group (Rubin et al. 2007), who recommended using spatially explicit capture recapture (SECR) methods on multiple small (2 x 6 trap) grids. Density and population estimates from annual trapping have been used to document recovery of listed Island Fox populations (Coonan et al. 2010a).
Recovery tool
Results from population and mortality monitoring have been incorporated into a “recovery tool” for land managers to use to assess recovery status of listed fox subspecies. Recent demographic modelling incorporated life-history characteristics of the well-studied Island Fox with environmental drivers and uncertainty to develop extinction probabilities for combinations of population size and annual mortality (Bakker et al. 2009, Bakker and Doak 2009). The demographic traits that best predicted extinction risk were adult population size and adult annual mortality rate, assessed jointly (Bakker and Doak 2009). Subspecies-specific graphs were developed that plotted risk isoclines across range of three-year average adult population sizes and mortality rates. Three-year averages of both adult population size and adult mortality have become the basis for assessing demographic recovery criteria for the Island Fox. Island managers and the Island Fox working group identified a predicted risk of quasi-extinction (decrease to a population size of <30) of less than five percent over 50 years as the population condition consistent with recovery. To account for uncertainty, the joint 80 percent confidence intervals for both population size and mortality must lie within the 5 percent extinction risk isoclines. Island Fox subspecies must meet this standard for five consecutive years before they may be considered for de-listing. This standard has now been adopted by the USFWS in the draft recovery plan for the Island Fox (USFWS 2012) as the demographic recovery criterion. As of 2012, use of the recovery tool indicated that San Miguel, Santa Cruz and Santa Catalina Island Foxes were close to demonstrating biological recovery, and Santa Rosa Island Foxes were several years from doing so (Coonan 2012).
Periodic serology
Island Foxes from all islands are periodically surveyed for evidence of disease exposure, as result of recommendations from the Island Fox working group regarding vaccination and serological testing. The Fox Health group recommended that blood samples from Island Fox populations be collected annually, and that such samples tested at least once every five years for evidence of exposure to CDV, canine parvovirus (CPV), canine adenovirus (CAV) and Toxoplasma. Island Fox populations had been tested previously for disease (Garcelon et al. 1992, Roemer et al. 2001, Clifford et al. 2006), and recent serological testing (Coonan 2012) confirmed earlier conclusions that each Island Fox subspecies has had a unique history of pathogen exposure, and vulnerability to various canid diseases is not equivalent among subspecies.
Threat mitigation plans
The USFWS’ draft Island Fox recovery plan requires threat mitigation plans to be developed by each land management agency before Island Fox subspecies can be delisted. Such plans must demonstrate the agency’s capability and willingness to detect and mitigate threats posed by the two most onerous mortality factors, predation and disease. An epidemic response plan has been developed for San Clemente Island Foxes (Hudgens et al. 2011) and plans are currently being developed for island foxes on the northern Channel Islands and Santa Catalina Island (B. Hudgens, IWS, unpubl. data). The plans detail appropriate pathogen monitoring and disease-specific mitigation actions, incorporating the specific operational capabilities of the respective agencies.
Gaps in knowledge
Density dependence
In some areas of San Nicolas Island, foxes have remained at a stable population density greater than 25 foxes/km2; however, this very high density is unusual in carnivore populations. Additional research on habitat use, diet and small-scale resource use, abundance, and fecundity are needed to explore density dependence and estimate carrying capacity on other Channel Islands. Research exploring the ecological drivers of the fox’s home range size, which varies significantly between studies, could have applications for disease transmission rates and inter-species competition, and ultimately inform us on density dependence by habitat type.
Meso-predator competition
Many of the islands contain meso-predator species that likely compete with the Island Fox. San Clemente, Santa Catalina and until recently, San Nicolas contained non-native feral cats. The removal of feral cats from San Nicolas Island in 2011 presents a unique opportunity to document the effects of competitor removal from an isolated and well-documented system. Santa Rosa and Santa Cruz Island contain the only native meso-carnivore, the Island Spotted Skunk (Spilogale gracilis amphiala), which likely competes with the Island Fox (Crooks and Van Vuren 1995, Roemer et al. 2002). Foxes are competitively dominant to skunks (Roemer et al. 2002), and the latter increased on both islands when foxes declined. Skunks have apparently declined on Santa Cruz Island during the recent recovery of foxes, and have not declined on Santa Rosa, where the recovering fox population is perhaps one-half of its pre-decline number (Coonan and Guglielmino 2012). Further refinement knowledge is required about the relationship between these two meso-predators.
Disease
Apart from the threat of non-native predators, the greatest threat on these conservation islands is the possibility of a mainland disease introduction. A number of recent studies have addressed the threat of disease, including previous pathogen exposure (Clifford et al. 2006), pathogen transmission (Sanchez 2012), and Canine distemper virus titer persistence. However, additional research and management may be needed to prevent outbreaks, as new transmittable diseases arrive on the California coast.

